
Prof. dr Walter Schlegel
Odczyt wygłoszony 2.10.1987 roku z okazji 75-lecia SVŐ (fragmenty)
Wielce Szanowny Panie Przewodniczący !
Szanowni Państwo!
Aby mówić o szansach i niebezpieczeństwach inbredu należy najpierw zdefiniować, co jest inbredem. A w ogóle, przede wszystkim określić, dlaczego prowadzimy hodowlę, jakie są dziedziczenia cech, jakie istnieją możliwości i instrumenty prowadzonej z sukcesem hodowli. Obiecuję nie stosować tu matematycznych formułek, a jedynie wykazać, że hodowla jest zajęciem przewidywalnym, którego wyniki dają się udowodnić w sensie reguł zbudowanych na statystyce. W życiu można dyskutować o wszystkim, mieć najróżniejsze zapatrywania, jeśli jednak jakieś twierdzenie sprawdzalne jest metodą naukową (jest ogromna ilość takich metod), wówczas staje ono poza wszelką dyskusją, należy je przemyśleć i przyjąć.
Prowadząc hodowlę myślimy w kategoriach pokoleń, w tym wypadku pokoleń psa. Minęło 15.000 do 20.000 lat od czasu, gdy pies został udomowiony, a więc kiedy to wilk stał się zwierzęciem domowym. Przez miliony lat naturalnym hodowcą był sam wilk. Człowiek natomiast doprowadził do ogromnego zróżnicowania psów. Brał po prostu do krzyżówek te lub inne osobniki, myśląc tylko o tym, które mu pasują do aktualnych zadań. Postępował w myśl zasady: ci oto rodzice będą się nadawać najlepiej i dadzą najlepsze potomstwo. Jeszcze 100 lat temu kierunek hodowli zależał wyłącznie od widzimisię osób rzekomo bardziej doświadczonych. O ostatecznych sukcesach lub porażkach hodowlanych decydowała siła indywidualnych cech osobnika uprzywilejowanego w takiej selekcji. W następnej kolejności zaczęto zastanawiać się nad dziedziczeniem cech. Według Mendla występują pośrednie procesy dziedziczenia (dwie różne predyspozycje ze strony matki i ojca prowadzą do powstania u potomstwa cech pośrednich). Istnieje dominująco recesywny proces dziedziczenia cech (recesywny allel uzewnętrznia się tylko wówczas, gdy występuje w podwójnej ilości, a więc jest homozygotą. Dziedziczony bywa jednak tak samo często, jak allel dominujący, który uzewnętrznia się również jako heterozygota).
Wchodzimy tu wówczas w temat inbredu, jego niebezpieczeństw i szans. Jeśli wezmą Państwo pod uwagę pulę genową, czyli ogół zawartych w genach danej populacji jej cech dziedzicznych, np. owczarków w Austrii, w Europie, na świecie, wówczas istnieje około 150 000 locci u każdego pojedyńczego psa (loci – niem. Genort, miejsca zajmowane przez geny tej samej cechy w chromosomach – przyp. tłum.). Z tej ilości co najmniej 1/3, (a prawdopodobnie trochę więcej), jest polimorficzna, tzn. w tym pojedyńczym locci są co najmniej dwa warianty przekazu cech, czasem 5 lub 6, a czasem 1000. Z tych wariantów potomkowi zostanie przekazany tylko jeden ze strony ojca i tylko jeden ze strony matki.
Inbred oznacza redukcję genetycznej różnorodności i tym samym homogeniczność, a więc ujednolicenie wyników hodowlanych tak, by cel, do którego się zmierza, był możliwie homozygotyczny. Ale uwaga: każdy kierunek hodowli, czy na umaszczenie, wielkość, charakter, witalność, kondycję jest zarazem zależny od istniejącej w danym osobniku puli genowej, a im wiecej locci jest homozygotami, tym zwierzę jest mniej sprawne. Spostrzeżenie to poczyniono niecałe 10 lat temu i zrewolucjonizowało ono myślenie wielu genetyków.
Jay Lush w swojej książce z roku 1938 „Animal breeding plan” opisał hodowlę większych populacji zwierząt i roślin. Stwierdził, że sukcesy w selekcji można przewidzieć. Doszedł też do niespodzianek, takiej na przykład, że heterozygotyczne zwierzęta okazały się nagle lepsze od tych, które były hodowane „na linię” i właściwy inbred.
Musimy zatem zadać pytanie, co tak na prawdę wpływa w populacji hodowlanej na pulę genową? Najpierw oczywiście mutacja. Mutacje stanowią zmiany w genotypie, które nie były przewidziane, nastąpiły spontanicznie i przebiegały bez określonego kierunku. Ale u nas mutacje ważne, brzemienne w konsekwencje, dzieją się stosunkowo rzadko. Są swoistą wymówką tylko wówczas, gdy coś zupełnie się nie uda. Wtedy mówi się z żalem o mutacji. Jest ona trudna do udowodnienia, bo należałoby ją dopiero sprawdzać poprzez doświadczenia na zwierzętach i poprzez świadome i planowane działanie wykorzystać zaistnienie takiej mutacji. Wyrzucam więc mutację poza nawias.
Weźmy migrację, czyli włączenie takich nowych cech dziedzicznych do populacji, których dotąd tam w ogóle nie było. Mamy z nią do czynienia wówczas, gdy np. wprowadza się zwierzęta hodowlane z zagranicznych populacji.
Pozostaje zatem selekcja! Zakłada ona jeden cel, niezależnie od tego, na co nastawiona jest hodowla, a wynik selekcji ma być osiągnięty w ciągu roku. Należy najpierw ustalić intensywność selekcji, tzn. ilu reproduktorów użyjemy w hodowli. Określamy także interwał pokoleniowy, tzn. przyjmujemy średnią jego wartość dla obojga rodziców na moment narodzin potomka.
Gdy jednak zastosujemy zbyt małą ilość par rodzicielskich, to dojdzie niezaprzeczalnie do genetycznego dryfu. Dryf taki jest fenomenem i zdarza się zawsze i u wszystkich gatunków zwierząt przy zbyt małej bazie hodowlanej. Genetyczny dryf oznacza dla populacji powstanie konsekwencji inbredu bez stosowania inbredu! Można dokładnie wyliczyć ile wystąpi tych konsekwencji mimo braku w działaniu znamion typowego inbredu!

Rozpatrzmy taki fakt: w małej grupie hodowlanej jednej rasy, np. bullterierach, otrzymujemy tylko 10 miotów z 2 lub 3 reproduktorów. Wówczas, nawet jeśli wszystkie zwierzęta nie są wcale ze sobą spokrewnione, możemy określić wzrost inbredu całej populacji w tym pokoleniu, którego wynik wyliczamy. Dzieje się to następująco: 1 dzielone przez 8 razy liczbę użytych w roku reproduktorów plus 1 dzielone przez 8 razy liczbę użytych w roku suk.
Gdy były to tylko 2 reproduktory, to jest to jedna/szesnasta (bo 1 / [8x2] ), a to już jest ogromnie dużo, bo to oznacza przecież już ponad 6% inbredu (jedna/szesnasta odpowiada 6,25% inbredu), a my nie powinniśmy nigdy przekraczać przeciętnie 6% dla całej populacji. A teraz dodajemy jeszcze liczbę wynikającą z jednej/osiemdziesiatej dla suk (bo 1 / [8x10] ), co daje 1/16 + 1/80 = 6/80; czyli 7,5% wzrostu inbredu.
I to wszystko w jednym pokoleniu! Ogólnie przyjmuje się, że przeciętny współczynnik inbredu w jakimkolwiek gatunku nie powinien przekraczać 6%. Te 6% obowiązuje dla nas homo sapiens jak i koni wyścigowych, krów, świń, muchówki, insektów i krokodylie.
O ponad 6% zmniejsza się wydolność zwierząt, a w konsekwencji rośnie ich ogólna ułomność. Występują inbredowe depresje, mimo że nie stosowano inbredu i w hodowli nie użyto do krzyżówek jakichkolwiek spokrewnionych ze sobą zwierząt!
A teraz kilka uwag o selekcji jako czynności hodowlanej. Selekcja, jaką my powinniśmy i chcemy prowadzić, wymaga posiadania dokładnych i wystarczających danych, które można wstawić do tzw. „krzywej Gaussa”. Należy wówczas wyliczyć odchylenie od standardu (δp)
i określić intensywność selekcji (i) oraz – jeśli znana jest odziedziczalność, którą oznaczymy jako h2 – zastosować odpowiednie działania hodowlane.
W obliczeniach tych T = interwał pokoleniowy (przeciętny wiek rodziców w momencie narodzin potomka przeznaczonego do dalszej hodowli), a t = rok.
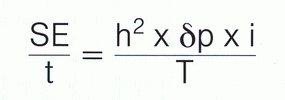
Przykład: przeciętna wysokość w kłębie psiej populacji wynosi 55,3 cm. Populacja ta ma zostać poddana selekcji w kierunku podniesienia tego wymiaru. Używa się w tym celu psów o największej wysokości w kłębie. Ich przeciętny wymiar wynosi 57,1 cm. Przy założonym współczynniku odziedziczalności (h) dla wysokości w kłębie wynoszącym 0,8 (poziom wysoki) daje się wyliczyć oczekiwany postęp hodowlany z pomnożenia 0,8 x 1,8 cm x 1. Otrzymany wynik 1,44 cm stanowi zamierzony sukces selekcyjny. Wówczas przeciętna wysokość w kłębie przyszłych pokoleń wynikać będzie z sumy przeciętnej wysokości w kłębie pokolenia rodziców i danego sukcesu selekcyjnego.
Ponieważ przeciętna wysokość w kłębie pokolenia rodziców wynosiła 55,3 cm.a sukces selekcyjny określiliśmy na 1,44 cm., więc oczekiwana przeciętna wysokość w kłębie przyszłego pokolenia wynosi: 55,3 x 1,44 = 56,74 cm.
W powyższy sposób, jeśli znamy odziedziczalność oraz wyjściowe i projektowane wartości selekcji, można wyliczać sukces selekcyjny dla każdej wymierzalnej cechy. Daje się on umiarkowanie podnosić poprzez zwiększanie intensywności selekcji. Oczywiście, gdy będą Państwo prowadzić hodowlę z użyciem najlepszych osobników, to osiągną sukcesy w selekcj. Można je wcześniej obliczyć nawet w skali roku także wówczas, gdy chodzi o poligeniczną cechę, np. charakter.
Większość istotnych cech będących celem hodowlanym nie jest kontrolowana jedynie przez pojedyńcze locci, nie są więc dziedziczone monogenicznie, ale wpływają na nie różne locci, znajdujące się w różnych chromosomach. Pojęcia takie, jak geny dominujące, wskazujące czy katalizatujące i inne pomijamy w tych rozważaiach. Obecnie przyjmuje się, że jest co najmniej 1 z przypuszczalnie 20 różnych współdziałających locci. Przy każdorazowym wzroście obciążeń genetycznych na locci wydolność organizmu zwierzęcego spada. Przy pomocy zdjęć rentgenowskich można w poziomach (stopniach) poklasyfikować skalę HD (dysplazji stawu biodrowego – przyp. tłum.). Następnie według niej ukierunkowuje się konkretne działania hodowlane. W tym miejscu należy wskazać na książkę "Hodowla psów w teorii i praktyce." (Walter Schlegel, Irene Stur: „Hundezuechtung in Theorie und Praxis; ein genetischer Leitfaden fuer erfolgreiche Rassehundezucht“, Verlag Jugend und Volk, Wien und München, 1986.)
Każde locci może zawierać jeden zdrowy allel (gen) i jeden chory allel (gen) albo dwa chore. Takie locci nie jest wówczas podwójnie chore, ale tak samo chore, jak w przypadku jednego zdrowego i jednego chorego allela. Tyle tylko, że zwierzę mające kilka locci podwójnie chorych (a więc od ojca i matki), będzie wcześniej przekazywać chorobę. I tu pojawia się konieczność, aby coś przeciwko temu uczynić. Na przykład porównywać potomstwo uzyskane po dobrych rodzicach. Jeśli zastosuje się właściwe środki zaradcze, wkrótce prawie zanikną osobniki o średniej do ciężkiej skali HD, natomiast częściej będą występować zwierzęta zdrowe, albo o lekkim lub granicznym obciążeniu.
Jeszcze raz wróćmy do monogenicznego, recesywnego dziedziczenia cech. Jeśli skrzyżują Państwo cechę rudości z czarną, to rudość jest cechą recesywną i wszystkie zwierzęta, które są heterozygotyczne będą wprawdzie czarne, ale z recesywną cechą rudości. Jeśli skrzyżuje się dwa takie zwierzęta, z których oba są czarne i heterozygotyczne, to potomstwo będzie w 3/4 czarne i w 1/4 rude. A z tych czarnych 1/3 będzie czarna – homozygotyczna.
Przy rudości i czerni ten często ukryty recesywny gen jest tylko kwestią umaszczenia. Przy recesywnych wadach genetycznych pochodzących z monogeniczności, w kategoriach populacji mamy często do czynienia z regularną bombą z opóźnionym zapłonem.

Niepożądany gen może mieć dla populacji straszliwe skutki. Weźmy przykład migracji takiego recesywnego allela wywołujacego wnętrostwo poprzez dokupienie wyglądającego zdrowo reproduktora, ale będącego nosicielem heterozygotycznym. Przy całkowicie zdrowej żeńskiej części populacji sytuacja ta oznacza, że gen ten wystąpi u 25% potomstwa. Mówimy wtedy, że częstotliwość występowania tego chorego recesywnego genu wynosi 25%, a zdrowego genu w postaci dominującej wynosi 75% (patrz rys nr 3).
Ale żaden z potomków nie jest jeszcze w sposób zauważalny chory. Jeśli używamy w dalszej hodowli nadal heterozygotycznych reproduktorów, to wówczas odsetek ten wynosi już prawie 40%, a w kolejnym pokoleniu 44%. I być może tu dopiero pojawi się osobnik, który będzie zauważalnie chory.
Fakt taki bywa z początku przemilczany. Rzecz zrozumiała. A wtedy w krótkim czasie mają Państwo w populacji wiecej chorych genów niż zdrowych, a usuniecie ich stanowi niesłychanie trudny problem. Jeśli na stałe wyłączyć z hodowli chore osobniki, to wprawdzie spada ilość chorych osobników homozygotycznych, ale heterozygotyczne w ilości 25% ciągle tam występują. Gdy z kolei poddamy selekcji rodziców tych osobników, to będzie trwało aż pełne 21 pokoleń, a więc około 60-70 lat, zanim częstotliwość występowania chorego genu spadnie z 10% do 0,1%. Ale to wcale nie jest okoliczność pożądana! Jako genetyk muszę stwierdzić, że powinniście być Państwo zadowoleni, iż macie w populacji obciążające, recesywne geny, bo to one w formie heterozygotycznej współdecydują o wydolności organizmu.
I tu przechodzimy konkretnie do postawionego zagadnienia.
Najpierw definicja wydolności organizmu (w tekście Fitness – przyp. tłum.) w największym skrócie: płodność, łatwość odchowu, długość życia, odporność na choroby, jednym słowem wszechstronna sprawność fizyczna organizmu! Potrzebujemy osiągnięć na najwyższym poziomie. Musimy zatem dążyć do tego, aby nasze zwierzęta były poddawane twardemu wychowaniu i mogły podejmować trudne zadania. Sprawność fizyczna oznacza bowiem również wytrzymałość, a szybkie męczenie się jest przecież jej zaprzeczeniem. Istnieją bardzo wyraźne związki między właściwościami składającymi się na wydolność organizmu a heterozygotycznością możliwie wielu locci całego genomu. Ponieważ powiedzieliśmy już, że około połowa loci ma możliwośc posiadania w sobie dwóch różnych alleli, to musimy zauważyć, że jeśli jeden osobnik jest w mniej niż 1/3 genów heterozygotą, tzn. zawsze będzie homozygotyczny, to jego wydolność będzie słabła, degenerowała się i wymierała.
Przykład: zwycięzcy wszystkich wielkich wyścigów konnych ostatnich 100 lat odbywających się corocznie w jednym miejscu, jak Pardubice, Arc de Triomphe, Derby itd., nie mieli nigdy współczynnika inbredu przekraczającego 2,5%. Przeciętnie było to 0,5%, a to oznacza, że byli właściwie „outcross”. Wszyscy mieli wysoki stopień heterozygotyczności, chociaż od pokoleń byli hodowani tylko na jedną cechę, mianowicie na sprawność ruchową.
Doświadczonym hodowcom nigdy nie przyjdzie do głowy prowadzić hodowlę na identycznych przodków, bo oznaczałoby to nieuchronnie zwiększenie inbredu.
W hodowli bydła doprowadziliśmy w ostatnim czasie przy pomocy wspaniałej metody „sztucznego zapłodnienia”, a może i „transferu embrionów” do tego, że coraz mniej byków względnie krów uczestniczy w akcie płodzenia potomstwa. Jedynym stawianym sobie celem hodowlanym jest mleczna wydajność, tyle tylko, że do wspomnianej wysokiej wydajności można dochodzić przez 5, 10, 12 lat przy płodności rzędu 5,10,12 cieląt. Można też uzyskać ją w ciągu 2-3 lat, używając najlepszych odżywek, w najlepszym środowisku naturalnym, stosując szczepienia i lekarstwa. Wówczas krowa może dawać rocznie 10-12 tys. litrów mleka, lecz nie urodzi już trzeciego potomka. Są kraje, gdzie prowadzono taką wysokokwalifikowaną, modelową hodowlę i poniosła ona tam kompletną porażkę. Przeciętna płodność karmiących potomstwo krów wyniosła tylko ponad 2,2 urodzin. Ponieważ co drugie narodziny przynoszą potomstwo żeńskie, więc każdy nowy osobnik żeński musi być włączony do populacji, aby zapewnić konieczną odbudowę stada.
Stosuje się obecnie, jakby w ostatnim ratunku, heterozygotię, ale to oznacza krzyżowanie ras! A tego w hodowli psów przecież nie chcemy! Ale czy aby na pewno?
Cunningham, doprawdy genialny genetyk z Dublina, przedstawił kilka lat temu tzw. wykres wydolności (patrz rys nr 4).

Można wstawić do niego wszystko, co ma do czynienia z wydolnością, a więc także cechy psów użytkowych. Im jakaś populacja leży na wykresie wyżej i po prawej stronie, tym ma wyższą wydolność i jest zarazem w wyższym stopniu heterozygotyczna. Natomiast znajdując się niżej i odchylając w lewo, jest wprawdzie homozygotyczna, mamy więcej psów z „doskonałych”, ale osiągi wydolnościowe spadają. Bardzo proste wyjaśnienie tego faktu dał nam Duńczyk Andresen (patrz rys nr 5).

Cała nasza przemiana materii, całe nasze życie, sterowane jest przez enzymy, gdyż to enzym przetwarza każdy substrat w życiodajny produkt. I tak dzieje się bez końca. Kiedy więc wyobrażą sobie Państwo, że ten enzym kontrolowany jest w znany Wam sposób genetycznie i każdy z nich jest homozygotą, A-A lub B-B, to wówczas może oddziaływać na substrat zawsze tylko jednym allelem i wytwarzać stale tylko taki sam produkt. A gdy ten produkt znowu jest przetwarzany, na przykład przez enzym B-B, to powstaje ponownie tylko jeden produkt. Tymczasem, gdy enzymy są heterozygotą (A-B), nieustannie powstają dwa różne produkty służące organizmowi. Oznacza to, że taki osobnik lepiej zaadaptuje się w środowisku, w którym przyjdzie mu żyć. Lepiej zniesie stres, niedożywienie, przepracowanie, niż osobnik, który jest homozygotyczny.
Gdy zdamy sobie sprawę, że na ogół w sumie jest 15-30 takich obszarów oddziaływania enzymów w sekwencji prowadzącej do końcowego produktu, to można pojąć, jak gigantyczne różnice istnieją w wydolności między zwierzętami z tych samych warunków hodowlano-środowiskowych i to tylko dlatego, że jedno jest możliwie często heterozygotyczne, a dla innego wspomniana sekwencja przynosi efekt homozygotyczny. Ale jakże by inaczej, skoro organizm tego pierwszego jest w stanie wybierać między 120 a 180 możliwymi reakcjami obronnymi, a ten drugi ma do dyspozycji zaledwie 20-30 mechanizmów obronnych. Wnioski te dotyczą wszystkich gatunków zwierząt, a więc odnoszą się i do człowieka.
Przejdźmy do kwestii wad genetycznych i czynników śmiertelności. Dotąd znanych jest około 1100 wad genetycznych psa, u człowieka znamy ich już około 1380 i co najmniej 1/3 podlega omówionym wcześniej mechanizmom. Dotyczą one stale związków między wydolnością organizmu a częstotliwością zmian w rozmieszczeniu genów, więc również zastosowanych metod hodowlanych, w tym inbredu.
W ten sposób wykazalismy w wystarczającym stopniu niebezpieczeństwa płynące ze stosowania inbredu. W hodowli owczarka nie stosuje się inbredu, co najwyżej chów na linię.
Zalecałbym jednak, także osobom odpowiedzialnym za hodowlę owczarka, przemyśleć pokazane wyżej analizy i stopniowo zwiększać liczbę używanych reproduktorów. Przy 32.000 sztuk szczeniąt rocznie wydaje się, że skutków inbredu nie potrzeba się obawiać.
Ale nie można przeoczyć faktu, że z tych 32.000 tylko niewiele będzie użytych do dalszej hodowli. Połowa osobników obu płci wykorzystanych w przyszłości do hodowli pochodzi z około 1% użytych reproduktorów! Tym samym ten ogromny potencjał wyjściowy zawęża się do konkretnych zdarzeń selekcyjnych, a wówczas liczby te nie są już takie wolne od zastrzeżeń!
Rasa ta musi dalej istnieć i jak długo będę żyć nie przestanę ostrzegać. Ciągle jeszcze można mówić o wysokiej jej płodności. Ale kiedyś rejestrowano 13 i 14-krotne mioty, co wymagało przecież upartej selekcji. Dziś stwierdza się całą masę miotów pojedynczych i podwójnych i podkreślam ten fakt z całym naciskiem. Nie są to jeszcze wyniki apokaliptyczne, ale ostrzeżenie, które należy zauważać. Baza hodowlana i liczba zdrowych linii jest przecież wystarczająco duża, a poprzez przemyślane krzyżówki wkrótce zbudować można nowe.
Bardzo dziękuję Państwu za uwagę!
Tłumaczenie – B. Demby
Korekta – I. Lelonek
Luty 2010

